 This is an open access article distributed under the terms of the Creative Commons
Attribution License (
This is an open access article distributed under the terms of the Creative Commons
Attribution License (Introduction
The basic mechanisms supporting spatial ability have received a great deal of attention from experimental psychologists (Nyborg, 1983; Smith & Chatterjee, 2008; Tommasi, Chiandetti, Pecchia, Sovrano, & Vallortigara, 2012). Amongst their many findings, consistent gender differences in tasks requiring manipulations and transformations of geometric figures and forms like mental rotation tasks, remain one of the most intriguing (for reviews, see Coluccia & Louse, 2004; Linn & Petersen, 1985; Voyer, Postma, Brake, & Imperato-McGinley, 2007; Voyer, Voyer, & Bryden, 1995). When considering the potential factors thought to support gender differences in spatial abilities, including mental rotation tasks, a recursive general hypothesis suggests that they are the result of differences in the strategies used to process information. However, since strategy differences are directly related to underlying cognitive functions, we define here the various cognitive functions thought to influence performance in mental rotation tasks.
Performance on mental rotation tasks may be influenced by gender differences in information processing. Mental rotation tasks based on comparison – where participants are asked to decide whether a pair of objects are identical or not – show consistent performance differences in favour of men. Findings from these kinds of tasks are the basis for the hypothesis that men employ a holistic, global approach to mental rotation in which they rotate the entire stimulus as a whole and then compare it to the target, whereas women employ a local, piecemeal approach in which individual features or parts are rotated and compared separately, piece by piece, to the target (e.g., Heil & Jansen-Osmann, 2008; Kail, Carter, & Pellegrino, 1979; Rilea, 2008).
Investigations varying the number of figures that subjects need to match have provided additional data regarding the different strategies that men and women use. These studies suggest that men rely more on a match-jump strategy: for example, once men have identified the matching target, they jump to the next problem without verifying the remaining stimuli, which presumably do not match, and thus progress through the test more quickly and have a chance to respond to more questions. In contrast, women tend to match every single figure to the target, even after identifying a match, and thus do not advance through the test as quickly, and often are unable to finish tests that have a fixed time limit (e.g., Glück & Fabrizii, 2010; Hirnstein, Bayer, & Hausmann, 2009).
In addition to a slower speed of responding, examination of the relations between self-reported confidence, performance and guessing behavior on mental rotation tasks indicates that men have a more accurate perception of their performance and were significantly more confident in their responses than women (e.g., Blough & Slavin, 1987; Cooke-Simpson & Voyer, 2007; Delgado & Prieto, 1996; Goldstein, Haldane, & Mitchell, 1990). This difference in confidence between men and women was similarly observed using self-assessment questionnaires of orientation skills (Coluccia & Louse, 2004; Schmitz, 1997), as well as in a study of exploratory behaviour (Brandner, 2007).
Performance on mental rotation tasks may be influenced by differences in the reference frame used by men and women. The differences between men’s and women’s use of global vs. local strategies have also been shown in experimental studies of orientation and navigation. For example, studies have shown that men’s use of a Euclidean strategy improved their performance (e.g., Astur, Tropp, Sava, Constable, & Markus, 2004; Moffat, Hampson, & Hatzipantelis, 1998; Sandstrom, Kaufman, & Huettel, 1998) whereas positional or landmarks strategies improved the performance of women (e.g., Astur et al., 2004; Levy, Astur, & Frick, 2005; Saucier et al., 2002). Further examination of the correlations among spatial abilities has revealed a significant relationship between mental rotation ability and the ability to use Euclidean information. However no similar significant relationship has been observed with the ability to use landmark information (Astur et al., 2004; Saucier et al., 2002).
Performance on mental rotation tasks may be influenced by differences in spatial perception. If men do employ a global strategy based on Euclidean information, then the difference between sexes might result from differences in the perception of the vertical and horizontal axes of the figures. It has been shown that, compared with men, women are worse at judging where the water level should come to rest in a titled container, and less good at judging the gravitational vertical position of a rod inside a tilted frame (e.g., Linn & Petersen, 1985; Robert & Ohlmann, 1994). The perceptual influence of coordinate axes was also shown in a paper and pencil task that required estimating the position and orientation of target lines by matching each to an identical line from a semicircular array of choices. The performance difference (approaching the magnitude of that observed on the mental rotations test) in favour of men has been correlated with the use of the vertical and horizontal axes of the page frame (Collaer & Nelson, 2002).
Finally, performance on mental rotation tasks may be influenced by gender differences in memory. If women do indeed employ a local approach in which individual features or parts are rotated and compared separately, then the differences between sexes might result from the differing amounts of information necessary to encode and hold in memory (i.e., memory load) when using global versus local strategies.
It has been argued that better spatial performance in men (as compared to women) is due to a larger visuospatial working memory (Coluccia & Louse, 2004 for an overview). Although, depending on the study, researchers have found sex differences in favour of both women and men in spatial working memory (e.g., Duff & Hampson, 2001; Vecchi & Girelli, 1998), these studies did not show a specific relationship between working memory and spatial ability.
Other investigations suggested that a small but significant female advantage may exist for general episodic memory (e.g., Andreano & Cahill, 2009; Herlitz, Nilsson, & Bäckman, 1997; Herlitz & Rehnman, 2008). For instance, episodic memory has been shown to vary as a function of gender and the type of material to be remembered. Women performed at a higher level than men when remembering words, objects, faces, and object positions while men outperformed women when remembering routes both with and without environmental information (Herlitz & Rehnman, 2008).
Although these spatial tasks support the general hypothesis that gender differences in performance are the result of differences in the strategies used to process information, they are not equivalent and therefore probably do not assess the same abilities in the same way. Thereby, the difficulty to get a clear picture of what really differs with respect to spatial abilities between men and women remains. A key step might be to clarify how cognitive functions affect information processing, and thus influence strategies. One manner to approach the problem is to distinguish cognitive processes thought to be fundamental to spatial abilities (e.g., Usher & McClelland, 2001; Van Zandt, 2000; Yonelinas & Parks, 2007) and to test them systematically. To this end, we have developed a task that requires participants to discriminate between memorized targets and lure geometric patterns that had been rotated in a clockwise or counterclockwise manner. Gender differences in discrimination performance, response time and confidence rating were analysed. In addition, we used signal detection theory (SDT) to dissociate a sensitivity parameter (d-prime) involved in the discrimination process from a response criterion parameter involved in the decision-making process (c-bias) (Green & Swets, 1966). Finally, discriminant function analyses were used to determine the set of variables that allowed us the highest level of discrimination between the two genders.
Methods
Participants
Participants were 30 right-handed volunteers (15 males and 15 females) recruited from the campus of the University of Lausanne (Switzerland). The age range was between 18 and 30 years. After having obtained extensive study information, participants provided written informed consent prior to beginning the study. The study was approved by the local Ethics Committee, and the NIH guidelines for the use of human participants in research were respected.
Stimuli
An original 2-D visual stimuli was created by combining three black, two-dimensional geometric shapes (square, circle and hexagon) of about 13 mm2 each (Figure 1.A) using the E-prime software (version 1/2, Psychology Software Tools, Inc) and displayed on an IBM Pentium 100 MHz computer equipped with 16 MB of RAM and a 14-inch monitor. The square and the circle were aligned on the y-axis while the hexagon was placed 35° below the x-axis. Three additional target stimuli (S) were created by three 90° trigonometric rotations of the y-axis. These four stimuli (S1, S2, S3, S4) were used as target-preserved canonical references with both a vertical (S1 and S3) and a horizontal (S2 and S4) dimension (Figure 1.A).
Twenty-four additional non-target stimuli were created by counterclockwise (CCW) and clockwise (CW) rotations of the canonical references axes of each target stimulus according to three amplitudes (10°, 20°, 30°, Figure 1.B).
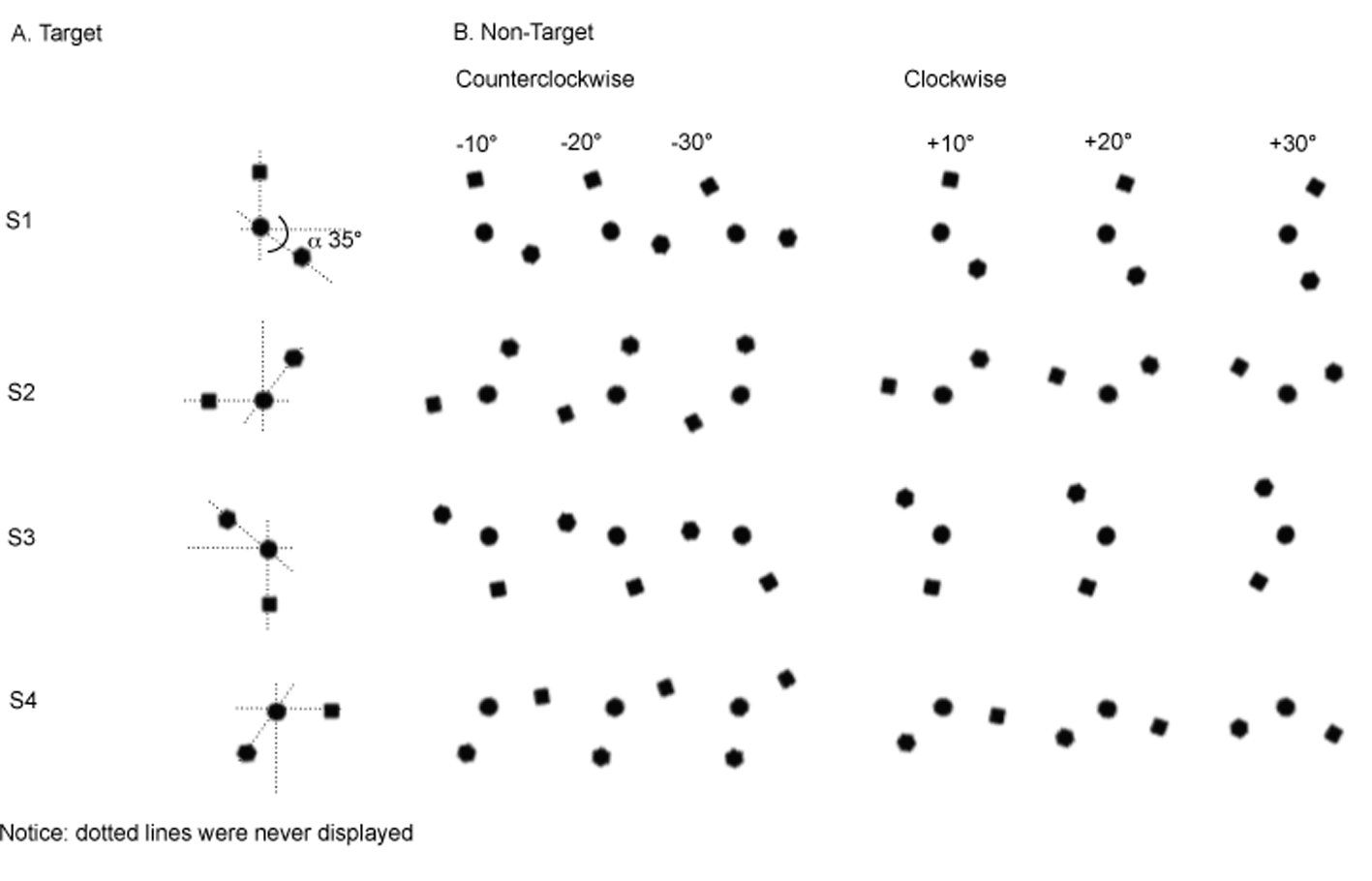
Figure 1
A. The 4 target stimuli of the learning phase. B. The 24 non-target stimuli of the discrimination phase.
Procedure
Instructions were given to each participant in written form displayed on the screen and were followed by 16 training trials with 4 similar targets but with different geometric shapes to familiarize the participants with the task. This training was followed by the learning phase where participants were asked to memorize each of the four target stimuli, displayed sequentially in the following order: S1, S2, S3, S4, presented for 5000 ms and followed by a blank screen for 3000 ms (Figure 2). The learning phase was immediately followed by a discrimination phase where participants attempted to discriminate as quickly and as accurately as possible, the targets from the rotated lure stimuli according to a "yes or no" forced-choice paradigm (yes = target; no = rotated lure). Each participant received 112 trials (16 target and 96 lure stimuli), and target and lure stimuli were randomly presented.
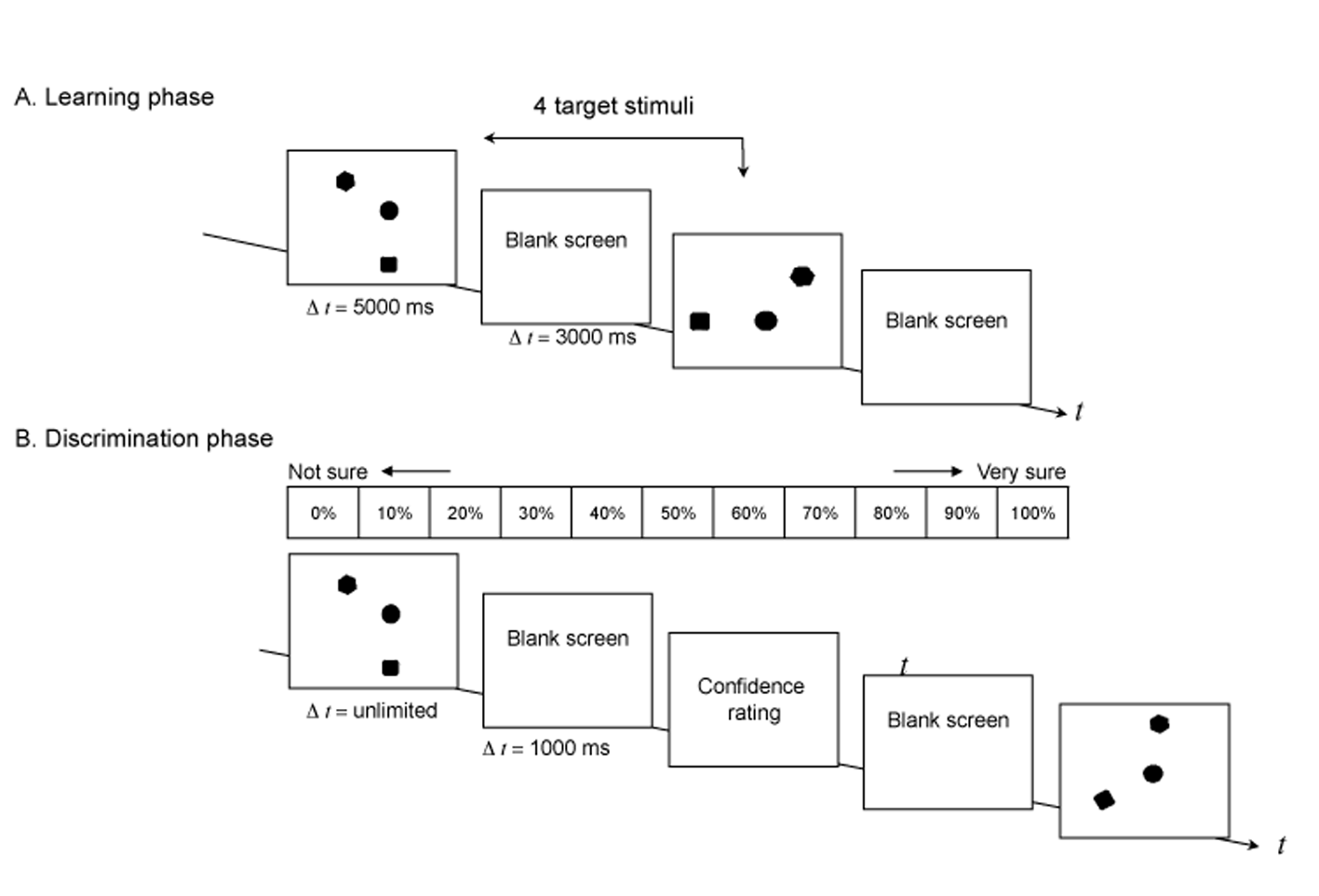
Figure 2
A. Outline of the learning phase. B. Outline of the discrimination phase.
The display remained visible until the participant responded and response was followed by a 1000 ms blank screen, which was then followed by a screen asking participants to assess their level of self-confidence (from 0% “do not know” to 100% “absolutely sure”) by clicking on the mouse with their right hand. After participants had rated their self-confidence, another blank screen would appear for 1000 ms, followed immediately by another stimulus presentation. Participants were not given any feedback regarding their performance during the task.
Results
Global Performance
A one-way ANOVA on the proportion of correct recognition revealed a statistically significant effect of gender (F [1, 28] = 4.62, p = .04; η2 = .14). As compared to women, men more often correctly identified target and lure stimuli (women: M = 0.74, SE = 0.04; men: M = 0.89, SE = 0.03; 95% CI [-0.29, -0.01]).
In contrast, neither the average confidence rating (F [1, 28] = 2.69, p = .11; women: M = 0.76, SE = 0.01; men: M = 0. 85, SE = 0.01; 95% CI [-20.17, -2.22]) nor the correct response time across responses (F [1, 28] = 1.98, p = .17; women: M = 2861.62, SE = 263.47; men: M = 2340.265, SE = 260.27; 95% CI [-237.26, 1279.98]) differed between men and women (Figure 3.A).
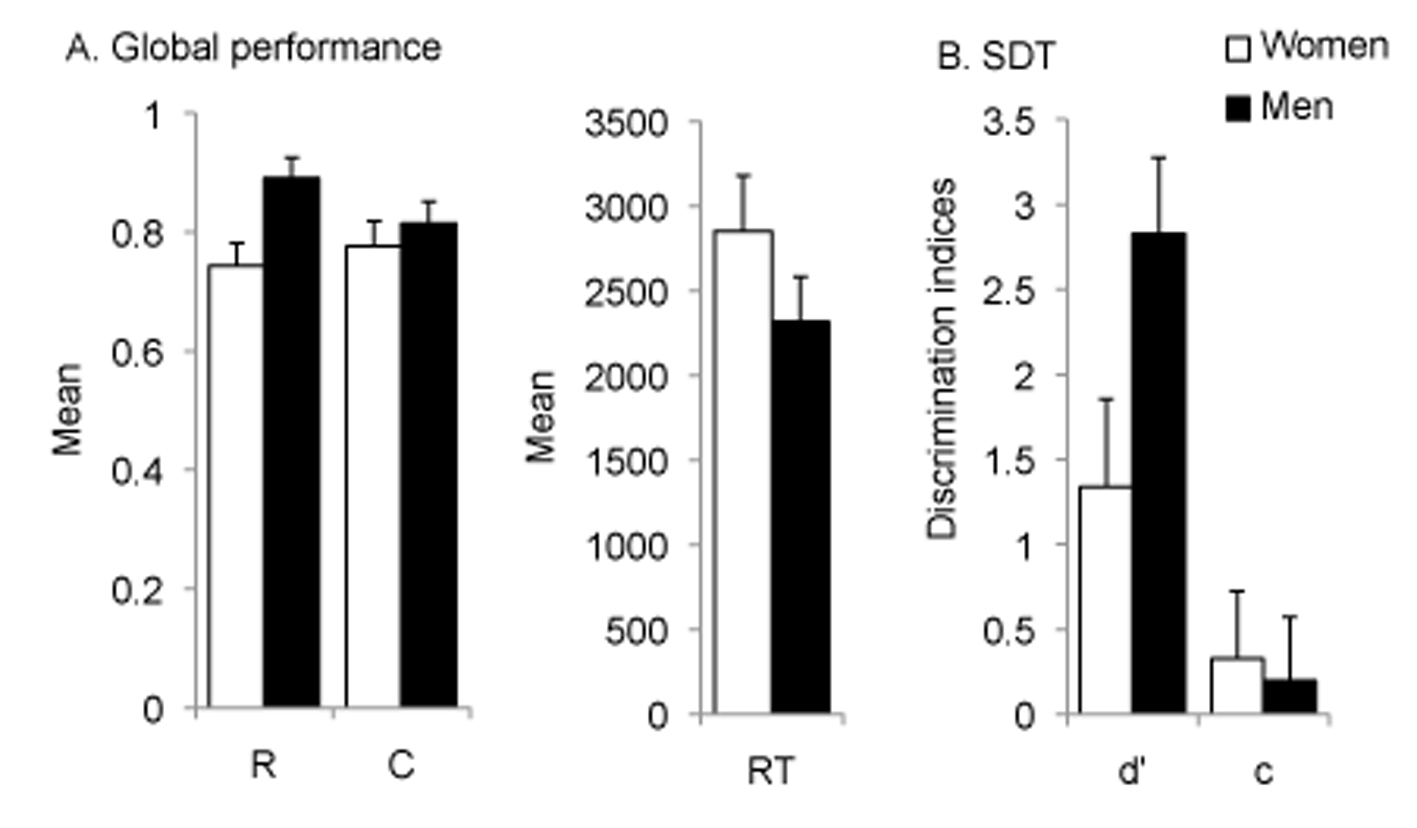
Figure 3
A. Correct recognition (R), confidence rating (C), and correct response time (RT) during the discrimination phase. B. Visual sensitivity and response criterion calculated on the full set of stimuli.
A multivariate ANOVA on d-prime and c-bias calculated on the full set of stimuli (Figure 3.B) revealed a higher d-prime for men as compared to women (F [1, 28] = 8.42, p = .01, η2 = .23; men: M = .28, SE = .13; women: M = .13, SE = .14; 95% CI [-2.54, -0.44]) while no difference between men and women was observed in c-bias (F [1, 28] = .75, p = .39; men: M = .19, SE = .34; women: M = .33, SE = .48; 95% CI [-0.18, 0.44]).
Assessment of Gender Differences as a Function of Target- and Lure-Specific Performance
As our task differed from the traditional tasks of mental rotation because it required both reference memory (learning phase) and working memory (discrimination phase), the responses of participants to target and lure stimuli were analyzed separately.
Target Stimuli (S1, S2, S3 and S4)
The averages for correct recognition, confidence rating and correct response time across responses for each target stimuli, S1, S2, S3 and S4, were respectively subjected to 2-way repeated measures (S1-S4) ANOVAs with a between–subject factor of “gender” and the mean of each of the dependent variables.
Across all target stimuli, recognition memory did not differ (F [3, 84] = .50, p = .69). Nevertheless, recognition of the target stimuli was statistically significantly better for men than for women (F [1, 28] = 9.17, p = .01 ; η2 = .25; 95% CI, [-0.45, -0.09]) while no statistically significant interaction between gender and target stimuli (F [3, 84] = .97, p = .41) was observed (Table 1).
Table 1
Correct Recognition (R), Confidence Rating (CR) and Correct Response Time (RT) by Women and Men for Each Target Stimuli (S1, S2, S3, S4)
| Mean
|
Standard error
|
|||||
|---|---|---|---|---|---|---|
| Target | R | CR | RT | R | CR | RT |
| Women (n = 15) | ||||||
| S1 | 0.60 | 0.77 | 3545.72 | 0.10 | 0.05 | 472.84 |
| S2 | 0.72 | 0.79 | 3016.69 | 0.09 | 0.05 | 248.30 |
| S3 | 0.53 | 0.77 | 2672.64 | 0.12 | 0.04 | 387.41 |
| S4 | 0.63 | 0.77 | 3317.99 | 0.10 | 0.04 | 536.34 |
| Men (n = 15) | ||||||
| S1 | 0.88 | 0.81 | 2203.40 | 0.07 | 0.04 | 339.10 |
| S2 | 0.88 | 0.85 | 2621.37 | 0.07 | 0.04 | 339.10 |
| S3 | 0.92 | 0.81 | 2139.75 | 0.05 | 0.04 | 292.70 |
| S4 | 0.88 | 0.80 | 2393.03 | 0.07 | 0.04 | 223.03 |
The same analysis on the average confidence rating (target stimuli: F [3, 84] = 1.94, p = .13; gender: F [1, 28] = .46, p = .50; target x gender: F [3, 84] = .28, p = .84) and on the average correct response time (target stimuli: F [3, 60] = .58, p = .63; gender: F [1, 20] = 2.78, p = .11; target x gender: F [3, 60] = 1.16, p = .34) revealed no other statistically significant difference (Table 1).
Lure Simuli
A 2-way ANOVA with multiple repeated measures (stimulus orientation S1-S4 x direction of rotation CW-CCW x amplitude of rotation 10°-30° x gender) for the average percentage of correct recognition revealed no statistically significant main effect (gender: F [1, 28] = 2.96, p = .10; stimulus orientation: F [3, 84] = .94, p = .42; direction of rotation: F [1, 28] = .01, p = .93; amplitude of rotation: F [2, 56] = .92, p = .40). This analysis did, however, detect a statistically significant interaction between gender and the direction of rotation (F [1, 28] = 5.95, p = .02, η2 = .18) as well as between gender, the stimulus orientation and the direction of rotation (F [3, 84] = 4.4, p = .01, η2 = .14). Pairwise comparisons of the interaction between gender and the direction of rotation revealed a male advantage over females in the correct recognition of lure stimuli that were rotated in a clockwise direction (F [1, 28] = 4.87, p = .04, η2 = .15; women: M = 0.74, SE = 0.05; men: M = 0. 91, SE = 0.05; 95% CI, [-0.32, -0.01]) whereas no difference between genders was detected in the counterclockwise direction of rotation (F [1, 28] = 1.31, p = .26; women: M = 0.78, SE = 0.05; men: M = 0. 87, SE = 0.05). Pairwise comparisons of the interaction between gender, the stimulus orientation and the direction of rotation revealed that women’s identification was poorer for the lure stimuli derived from Target Stimuli 1 and 2 and rotated in the clockwise direction (S1: F [1, 28] = 5.50, p = .03, η2 = .16; women: M = 0.78, SE = 0.07; men: M = 0. 94, SE = 0.03; 95% CI, [-0.32, -0.02]; S2: F [1, 28] = 5.90, p = .022, η2 = .17; women: M = 0.63, SE = 0.10; men: M = 0. 90, SE = 0.04; 95% CI, [-0.50, -0.04]) and for those derived from Target Stimuli 4 rotated in the counterclockwise direction (F [1, 28] = 4.52, p = .042, η2 = .14; women: M = 0.67, SE = 0.10; men: M = 0. 90, SE = 0.05; 95% CI, [-0.46, -0.01]).
The same analysis on the average confidence rating across responses revealed no statistically significant main effect of gender (F [1, 28] = 3.03, p = .09), stimulus orientation (F [3, 84] = 1.18, p = .32) or direction of rotation (F [1, 28] = 3.13, p = .09), but a statistically significant effect of amplitude of rotation (F [2, 56] = 4.98, p = .01, η2 = .15). Pairwise comparisons between the amplitudes of rotation revealed less confidence in the recognition of lure stimuli rotated by 10° as compared to lure stimuli rotated 20° and 30° (10° vs. 20° p = .012; 95% CI, [-4.07, -0.54]; 10° vs. 30° p = .041; 95% CI, [-4.02, -0.09], Table 2).
Table 2
Confidence Rating by Women and Men for Each Amplitude of Rotation
| Mean
|
Standard error
|
|||
|---|---|---|---|---|
| Amplitude | Women (n = 15) | Men (n = 15) | Women | Men |
| 10° | 0.74 | 0.84 | 0.04 | 0.04 |
| 20° | 0.77 | 0.86 | 0.04 | 0.04 |
| 30° | 0.76 | 0.86 | 0.04 | 0.04 |
This analysis did not detect statistically significant interactions between gender, stimulus orientation and direction of rotation.
The same analysis on the mean correct response time revealed no statistically significant main effect of gender (F [1, 21] = .41, p = .53), stimulus orientation (F [3, 63] = .91, p = .44), amplitude of rotation (F [2, 42] = 1.40, p = .26), or direction of rotation (F [1, 21] = 80, p = .38). This analysis did not detect statistically significant interactions between gender, stimulus orientation and direction of rotation.
A MANOVA on d-prime and c-bias calculated on clockwise and counterclockwise lures, respectively, revealed a statistically significant effect of gender for d-prime in both the clockwise condition (F [1, 28] = 12.00, p = .002, η2 = .30; women: M = 1.14, SE = .40; men: M = 2.82, SE = .281; 95% CI, [-2.67, -0.69]) and the counterclockwise (F [1, 28] = 6.19, p = .02, η2 = .18; women: M = 1.43, SE = .33; men M = .2.62, SE = .34; 95% CI, [-2.17, -0.21]) conditions. No gender effect was observed for c-bias in the clockwise condition (F [1, 28] = .03, p = .87, women: M = .22, SE = .13; men: M = .19, SE = .08) whereas a gender effect was nearly statistically significant for c-bias in the counterclockwise condition (F [1, 28] = 3.88, p = .06, η2 = .12; women: M = .37, SE = .11; men: M = .10, SE = .08; 95% CI, [-0.01, 0.55], Figure 4.A). Further paired comparisons between men and women revealed a statistically significant improvement in the women’s SDT parameters in the counterclockwise condition (d-prime/c-bias CW vs. CCW t(14) = -2.30, p = .04; 95% CI, [-0.57, -0.02]) due to a decrease in their false alarm rate (Figure 5.A) whereas no statistically significant difference was observed for men (d-prime/c-bias CW vs. CCW t(14) = 1.56, p = .14; 95% CI, [-0.07, 0.46]; Figure 4.B).
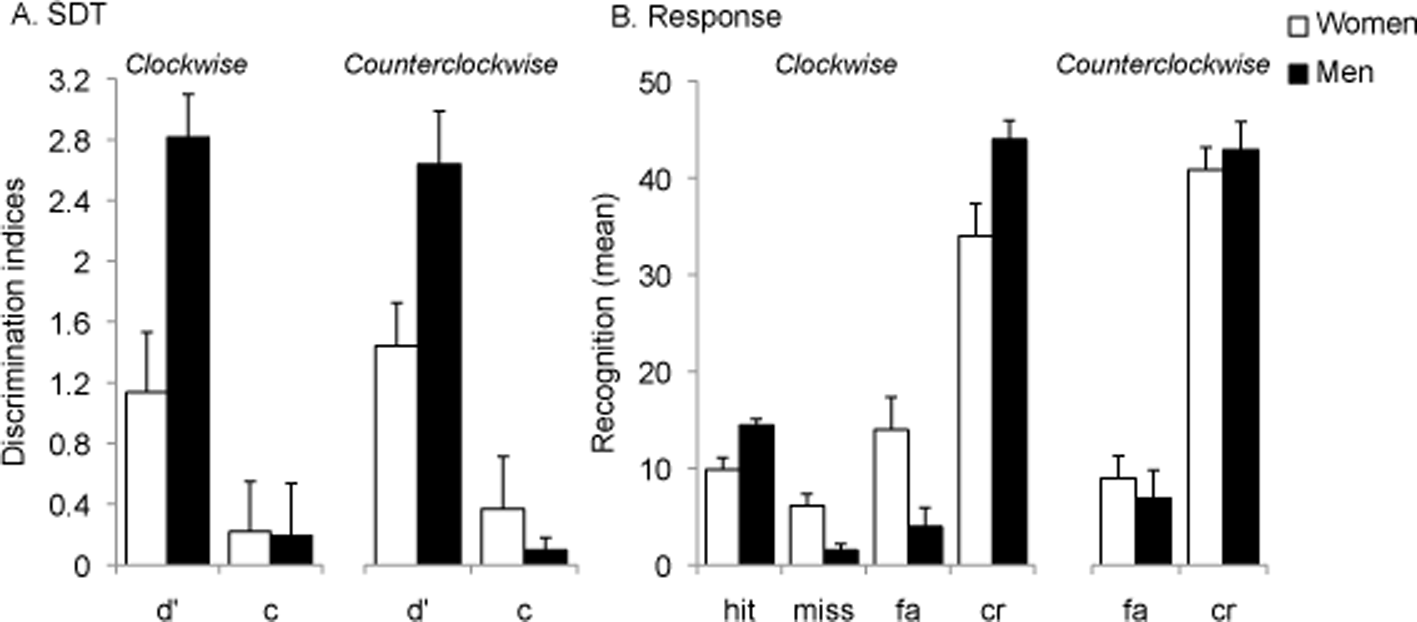
Figure 4
A. d-prime and c-bias values calculated on clockwise and counterclockwise subsets of stimuli. B. fa: false alarm; cr: correct rejection.
Discriminating Between Genders
Discriminant function analyses were used to determine the set of variables that allowed for the best discrimination between genders and also to evaluate the extent to which multiple predictor variables were related to group membership (i.e., women vs. men). They were performed on the full set of target stimuli, as well as on the subsets of clockwise (+10 °, +20 °, +30 °) and counterclockwise (-10 °, -20 °, -30 °) rotated lure stimuli, respectively. Predictor variables were d-prime, c-bias, recognition, confidence rating and correct response time. On the full set of stimuli, analyses showed significant mean differences for d-prime (F [1, 28] = 8.42, p = .08) and recognition (F [1, 28] = 4.64, p = .04) whereas no difference was observed for the remaining variables (c-bias: F [1, 28] = .75, p = .39; confidence rating: F [1, 28] = 2.70, p = .11; correct response time: F[1, 28] = 1.98, p = .18). Box’s M indicated that the assumption of equality of covariance matrices was respected (M: 24.97, F (15,3156.63) = 1.34, p = .17) despite the small sample. The discriminate function analysis revealed that the association between groups and all predictors failed to be statistically significant (χ2: 9.82, p = .08) and accounted for only 32% of the variability between men and women. This might be due to the fact that the d-prime score (.800) had the largest load of the function while the other variables were poor predictors. The cross validated classification showed that overall, 67% of the cases were correctly classified (Figure 5.A).
The same analysis performed on the subset of clockwise–rotated lure stimuli yielded similar results: (d-prime (F [1, 28] = 12.04, p = .002; recognition (F [1, 28] = 9.17, p = .005; c-bias: F [1, 28] = .30, p = .87; confidence rating: F [1, 28] = 3.48, p = .07; correct response time: F [1, 28] = .94, p = .34). The discriminate function analysis between groups and all predictors (χ2: 10.92, p = .05) was almost statistically significant, but accounted for 34% of the variability between men and women, and 73% of cases were correctly classified (Figure 5.B). Finally, the discriminate function analysis performed on the subset of counterclockwise-rotated lure stimuli revealed a statistically significant association between groups and all predictors (χ2: 12.95, p = .02), accounting for 40% of the variability between men and women.
This result was due to changes in the structure matrix where the significance of the d-prime predictor decreased (.58) and that of c-bias increased (-.46), whereas the other predictors remained weak. Accordingly, the correctly classified cases increased to 73% (Figure 5.B).
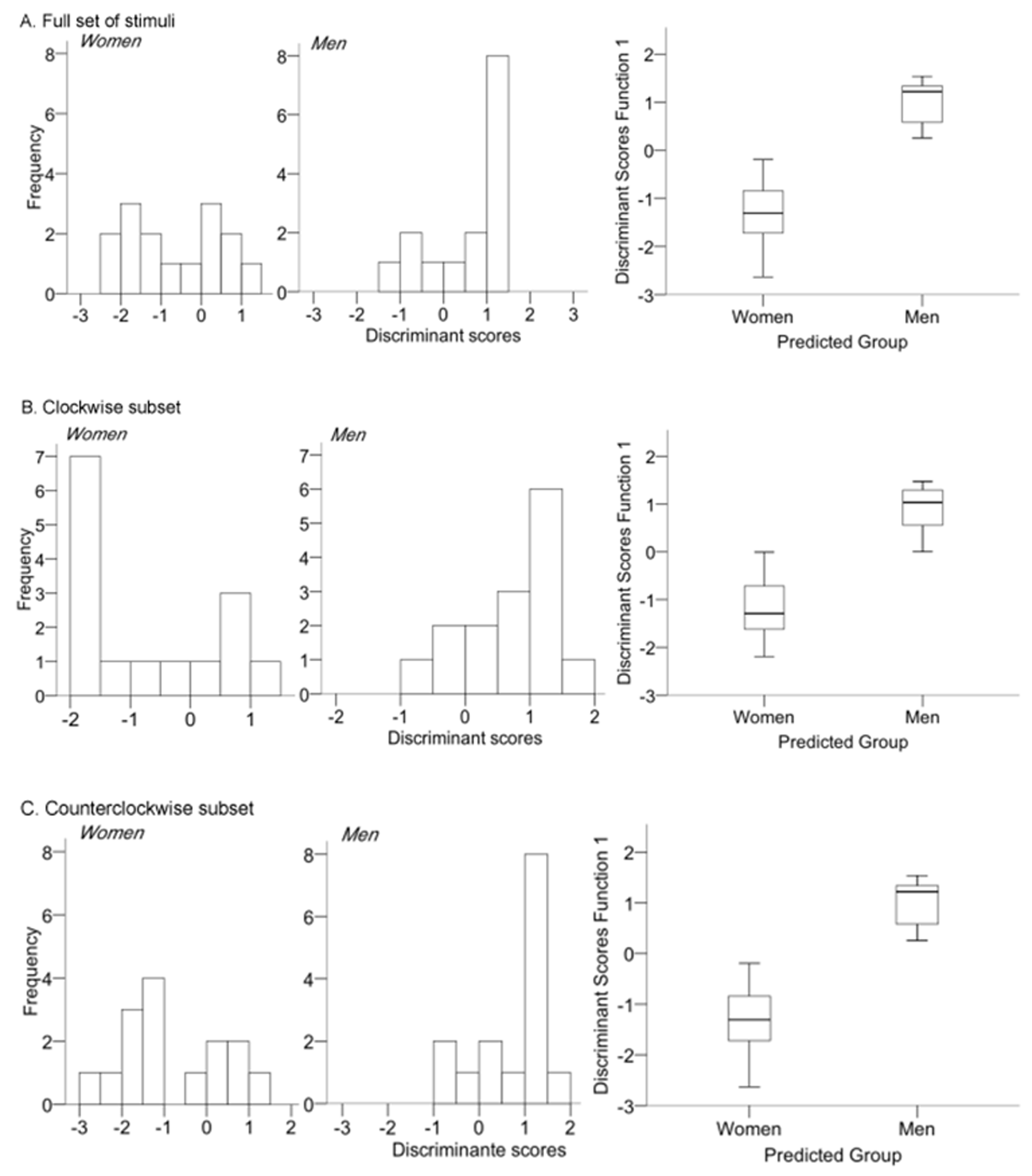
Figure 5
Distribution of the discriminant function scores and box plots for: A. Full set of stimuli; B. the clockwise and C. counterclockwise subsets of stimuli.
Experiment 2
Although one would assume that our task should be solved by performing a mental rotation of the target and/or lure stimuli, it is also possible that participants solve the task by simply noticing and remembering the alignment of the geometrical axes of the target stimuli. For example, all 4 target stimuli had the central circle and distal square aligned on either a vertical axis (i.e., at 0° for Target S1 and at 180° for S3) or a horizontal axis (i.e., at 270° for S2 and 90° for S4). The cardinal alignments of the target stimuli may be particularly salient as participants can compare the axes of the stimuli with the frame of the computer screen that is visible in all trials (Collaer & Nelson, 2002). Thus, in order to solve the task, participants only needed to have recognized that the target stimuli always had a circle and a square that aligned with one of the four cardinal axes, and then determine whether for the presented stimuli the square and circle aligned with a cardinal axis: If yes, the stimuli is a target stimulus; If no, the stimulus is a lure stimulus. We thus tested participants using a new boomerang-shaped stimulus as a target in order to further investigate this possibility (Figure 6).
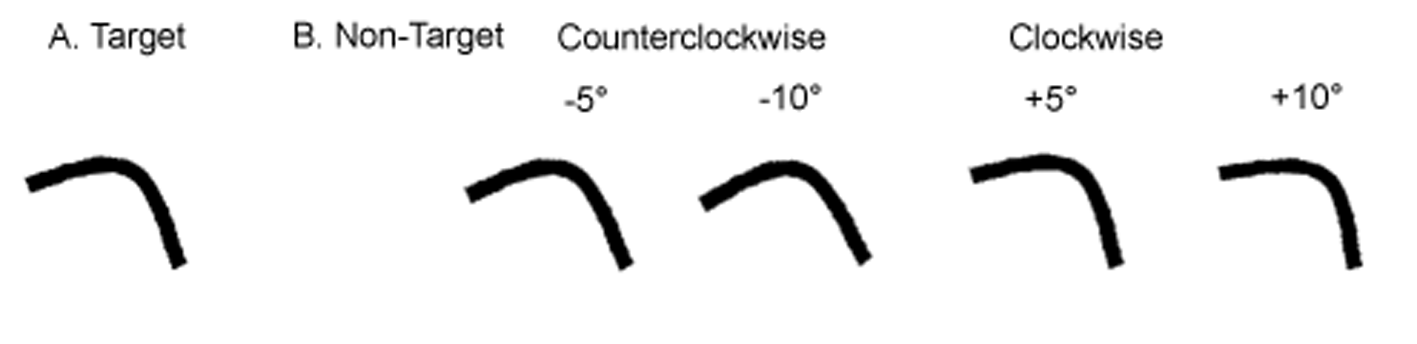
Figure 6
A. The 4 Boomerang-shaped target stimuli of the learning phase. B. The 24 non-target stimuli of the discrimination phase.
Importantly, the arms of the stimulus never aligned with cardinal axes. Non-target stimuli were created by counterclockwise (CCW) and clockwise (CW) rotations of this target stimulus at two different amplitudes (5° and 10°, Figure 2). Immediately following the testing with the four target stimuli (S1-S4) and their lures, participants were tested with the boomerang-shaped stimuli (Figure 6) following the exact same procedure. Each participant received 24 trials (8 target and 16 lure stimuli), and target and lure stimuli were randomly presented.
Boomerang-Shaped Stimuli
Global Performance
Separate one-way ANOVAs on the proportion of correct recognition (F [1, 28] = .06, p = .80; η2 = .002, women: M = 0.81, SE = 0.05; men: M = 0.79 SE = 0.07), the average confidence rating (F [1, 28] = .05, p = .83; η2 = .002; women: M = 0.72, SE = 0.03; men: M = 0.70, SE = 0.05) and the correct response time (F [1, 28] = .07, p = .79; η2 = .003, women: M = 2461,88, SE = 292.14; men: M = 2574.43, SE = 292.14) revealed no statistically significant effect of gender (Figure 7.A).
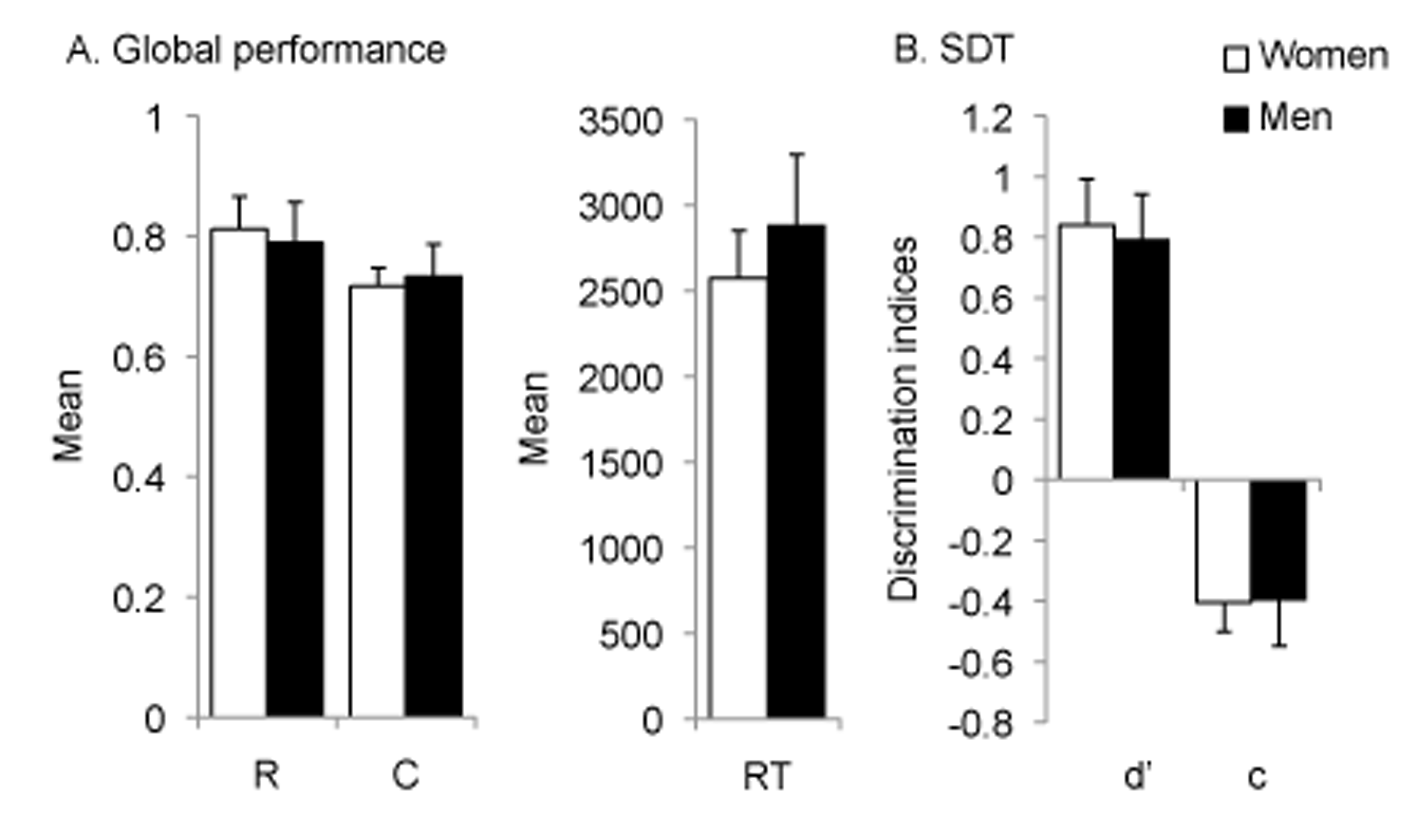
Figure 7
A. Correct recognition (R), confidence rating (C), and correct response time (RT) during the discrimination phase. B. Visual sensitivity and response criterion calculated on the full set of stimuli.
A multivariate ANOVA on d-prime and c-bias calculated on the full set of boomerang stimuli (Figure 7.B) revealed no effect of gender for d-prime (F [1, 28] = .05, p = .83, η2 = .002) or c-bias (F[1, 28] = .002, p = .97, η2 = .0001).
Lure Stimuli
A 2-way ANOVA with multiple repeated measures (direction of rotation CW-CCW x amplitude of rotation 5°-10°- x gender) for the average percentage of correct recognition revealed no statistically significant main effect of gender (F [1, 28] = .0001, p = .99) nor of direction of rotation (F [1, 28] = 1.64, p = .21) but a statistically significant main effect of amplitude of rotation (F [1, 28] = 25.70, p = .0001; η2 = .48; CW: M = .36, SE = .04; CCW: M = .65, SE = .04; 95% CI, [-.41, -.17]). This analysis did not detect statistically significant interactions between gender and the direction of rotation (F [1, 28] = .01, p = .92) nor the amplitude of rotation (F [1, 28] = .34, p = .56), but a statistically significant interaction between the direction of rotation and the amplitude of rotation (F [1, 28] = 4.7, p = .04, η2 = .14). Pairwise comparisons between the direction of rotation and the amplitude of rotation revealed higher levels of correct recognition when 10° lure stimuli were rotated clockwise than counterclockwise (5° CW vs. CCW: p = .60; 5° CW: M = .39, SE = .07; 5° CCW: M = .33, SE = .07; 95% CI, [-.16, -.27]; 10° CW vs. CCW: p = .08; 10° CW: M = .76, SE = .06; 10° CCW: M = .54, SE = .08; 95% CI, [-.03, .45]).
The same analyses on the average confidence rating across responses and on the mean correct response time revealed no statistically significant main effect and no statistically significant interaction of gender (confidence rating: sex F [1, 28] = .06, p = .81; rotation F [1, 28] = .00, p = 1. 00: amplitude F [1, 28] = 1.43, p = .24; correct response time: sex F [1, 3] = .991, p = .393; rotation F [1, 3] = .05, p = .83; amplitude F [1, 3] = .01, p = .91).
Discussion
The aim of this study was an attempt to synthesize and better understand gender differences in mental rotation tasks, specifically, and in overall spatial abilities, in general. To this end, data were analysed with classical statistical methods, but also with SDT parameters to dissociate perceptual from decision-making process. Finally, we assessed whether or not there are statistically significant multivariate differences between men and women on the specific variables of interest, including d-prime, c-bias, recognition memory, confidence rating and response time.
Global Performance
The average higher recognition performance in men, despite the fact that participants were given unlimited time to respond, is consistent with previous reports in mental rotation tasks (Coluccia & Louse, 2004 for reviews; Lawton & Morrin, 1999; Linn & Petersen, 1985; Voyer et al., 1995). In contrast to previous studies however, we found no differences between men and women in their overall confidence ratings or their correct response times (Cooke-Simpson & Voyer, 2007; Delgado & Prieto, 1996; Goldstein et al., 1990; Peters, 2005; Voyer, 1997; Voyer, Rodgers, & McCormick, 2004).
More sophisticated analyses (SDT parameters) of the processes involved in recognition showed that the male advantage in recognition memory resulted from higher visual sensitivity compared to women rather than a difference in the response criterion between men and women. These results indicate that, in this task, the differential performance between men and women is due to perceptual processes, whereas decision-making processes (c-bias) were equally biased by uncertainty. The results of our SDT analyses enable us to reject several hypotheses often evoked to explain gender differences observed in mental rotation tasks. First, the idea that women performed the task more carefully than men was not confirmed (Goldstein et al., 1990). The results of women and men were similar both with respect to confidence rating and response criterion. It is important to note that these two processes differ. Confidence rating was measured after each decision response (i.e., tapping the “yes” or “no” keys representing recognition of the presented stimulus as a target or not), whereas response criterion is calculated from each response and thus includes the process of selecting among several alternatives. Second, we can also reject the hypothesis that women, in general, take more time to solve the task and respond, which under limited time conditions, results in them guessing more often (Cooke-Simpson & Voyer, 2007). Guessing would bias responses and thus alter the response criterion of women, but our analyses did not show any difference in c-bias between men and women. Instead, our findings indicated that in this task, the differential performance of men and women was a result of differential visual sensitivity. This finding might be supported by others studies showing that, on average, men outperform women in spatial perception tests such as the Rod and Frame and water level tests that require participants to locate the gravitational upright despite the presence of distracting perceptual information (e.g., Rilea, 2008; Rilea, Roskos-Ewoldsen, & Boles, 2004; Tremblay, Elliott, & Starkes, 2004; for a meta-analysis see Voyer et al., 1995). However, just knowing that women exhibited poorer visual sensitivity does not allow us to determine whether this sensitivity varies according to the stimuli used. Further insight into this question might help to evaluate the hypothesis that the differential use of global and local strategies by men and women accounts for gender differences in mental rotation tasks (e.g., Heil & Jansen-Osmann, 2008).
Recognition of Target and Non-Target Stimuli
Although men exhibited a greater ability to recognize target stimuli that had been viewed during the learning phase than women, no gender difference was observed in the ability to recognize stimuli viewed only during the discrimination phase. This finding was unexpected. It has been argued that better spatial performance in men is due to a larger visuospatial working memory, whereas in women a small but significant advantage may exist for general episodic memory capacities (Andreano & Cahill, 2009; Herlitz et al., 1997; Herlitz & Rehnman, 2008). Reference memory is thought to correspond to the ability to recall or recognize the earlier-presented information while working memory is thought to require the simultaneous storage and processing of information. According to these definitions and the above hypotheses, recognition of the studied target stimuli, presumably implicating reference memory, should be relatively better in women, whereas recognition of non-studied lure stimuli, presumably implicating working memory, should be relatively better in men.
Further investigations including the other dependent variables manipulated in this study revealed other subtle differences between genders. Differences in recognition memory between men and women appeared to vary according to rotation direction. First, women exhibited poorer recognition for lure stimuli that were clockwise rotations of Targets 1 and 2 and for those that were counterclockwise rotations of Target 4 (see Figure 1). This unexpected result cannot be explained by our design because no difference was observed in the recognition of different target stimuli across genders. Thus, this difference cannot be due to particular characteristics of these target stimuli, for example, or to a serial effect of learning (Targets 1 and 2 were presented first and Target 4 last). Second, when boomerang-shaped stimuli were used (Experiment 2) participants showed a better rate of correct recognition for clockwise rotation than counterclockwise rotation, and when the amplitude of the rotation was 10° as compared to 5°.
Although effects of the direction of rotation are difficult to define, an EEG investigation of two potential strategies for determining the rotation of an object has shown that mental rotation of alphanumeric characters is more difficult in a counterclockwise than in a clockwise direction (Liesefeld & Zimmer, 2011). Although the effect of rotation could be related to task difficulty, it could also be related to differences in cognitive strategy. An earlier study provides a theoretical background to investigate this issue (Just & Carpenter, 1985). According to these authors, one strategy would be based on rotation of mental representations around canonical axes while the second one would rely on mental representations encoding spatial relationships between objects. Accordingly, the first strategy is not invariant with respect to the object’s orientation and position and thus involves the use of mental rotation to solve the task. By contrast, the second strategy would offer the advantage of being invariant with respect to the object’s orientation and position and thus allowing comparison or recognition without any mental rotation.
If participants had used a strategy consisting of mentally rotating displayed stimulus toward the canonical references (a vertical or horizontal virtual axes), the shortest way would entail rotating lure stimuli tilted clockwise in a counterclockwise direction, and rotating lure stimuli tilted counterclockwise in a clockwise direction. According to this hypothesis, stimuli rotated clockwise should be more difficult to recognize than those rotated counterclockwise. This is consistent with the results of our SDT analyses showing that, for women, counterclockwise rotations resulted in an improvement in their visual sensitivity due to a decrease in their false alarm rate. As making false alarms is due to a tendency to not discriminate between target and lure stimuli, the hypothesis would be that, for women, clockwise rotation is more difficult than counterclockwise rotation. In contrast for men, their ability to distinguish between lure and target stimuli (d-prime) was not altered by the direction of rotation. An alternative explanation, more consistent with previous research, would be that women rely on the first strategy of mentally rotating the displayed stimulus toward the canonical references of the memorized target until the two angles coincide. Accordingly, lure stimuli tilted clockwise are more difficult than those tilted counterclockwise.
Would this mean that men rely on the second strategy? The answer is probably no because if men were biased toward the second strategy, sex differences should be maintained when boomerang-shaped stimuli replaced geometric-shaped stimuli while they disappeared.
An explanation may be found by considering that, in general, men lend more attention to the geometry of the environment whereas women lend more attention to environmental landmarks. Because stimuli are displayed on a computer screen, perhaps men simply established whether the main axis of the displayed stimulus was aligned with the frame of the screen, indicating that the stimulus was a target. This suggestion is supported by an elegant study showing that when geometric cues (the page frame geometry) were eliminated, differences in performance between women and men disappeared (Collaer & Nelson, 2002). In accordance with this hypothesis, verbal reports from our participants at the end of the task indicated that men predominantly used this strategy, whereas women reported trying to recall stimuli from memory.
In conclusion, previous research suggests that women employ a local “piecemeal strategy”, while men rely more on a global one. Better spatial ability in men has been attributed to a greater visuospatial memory capacity despite its contradiction with local (involving larger amounts of information to encode) vs. global (involving a higher information compression) processing. The present results suggest that gender differences in strategies used to solve mental rotation tasks might be related to the ability of men to perceive and transform the problem into a simple task requiring pattern discrimination. Although, these results provide a base to tackle what really differs from the spatial abilities between men and women, they cannot yet be firmly validated because of the small sample size. Thus, further studies with larger sample sizes are needed.